Явление гетерозиса возникает при. Как называют явление гибридной силы. Аллельный вариант фермента
Гибридная сила heterosis, hybrid vigour - гетерозис, гибридная сила.
Превосходство гибридов по ряду признаков над родительскими особями, проявляющееся только в первом поколении и не передающееся по наследству; в основе Г . могут лежать более высокий уровень гетерозиготности <heterozygosis > гибрида, межаллельная комплементация <interallelic complementation > и др. причины; термин “Г .” предложен Г.Шеллом в 1911.
(Источник: «Англо-русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд-во ВНИРО, 1995 г.)
Смотреть что такое "гибридная сила" в других словарях:
гибридная сила
См. Гетерозис … Большой медицинский словарь
ГИБРИДНАЯ СИЛА - см. гетерозис … Словарь ботанических терминов
ГИБРИДНАЯ СИЛА - Конечный результат гетерозиса, т.е. фенотипическое превосходство гибрида над негибридными сверстниками … Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных
Гетерозис (или гибридная сила) - биологический эффект, связанный с превосходством гетерозигот над гомозиготами по степени экспрессии того или иного признака (эффект сверхдоминирования) и степени относительной приспособленности. Гетерозис создает основу для отбора в пользу… … Физическая Антропология. Иллюстрированный толковый словарь.
гибридная мощность - heterozė statusas T sritis augalininkystė apibrėžtis Padidėjęs pirmos kartos hibridų gyvybingumas (auga greičiau už tėvines formas, yra biologiškai produktyvesni, pajėgesni negu tėviniai individai). atitikmenys: angl. heterosis; hybrid vigour rus … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
гетерозис у растений - Ндп. гибридная сила Явление превосходства гибрида над лучшим из родителей по степени развития тех или иных признаков и свойств, в наибольшей степени проявляющееся в первом поколении. [ГОСТ 20081 74] Недопустимые, нерекомендуемые гибридная сила… … Справочник технического переводчика
- (от греч. heteroiosis изменение, превращение), «гибридная мощность», превосходство гибридов по ряду признаков и свойств над родительскими формами. Термин «Г.» предложен Дж. Шеллом в 1914. Как правило, Г. характерен для гибридов первого поколения … Биологический энциклопедический словарь
гетеродуплексный анализ г гибридизационный а - гетеродуплексный анализ, г. гибридизационный а. * гетэрадуплексны аналіз, г. гібрыдызацыйны а. * heteroduplex analysis or h. hybridization a. метод выявления генных мутаций путем смешивания амплифицированной при ПЦР мутантной молекулы ДНК с ДНК… … Генетика. Энциклопедический словарь
- (от греч. heteroiosis изменение, превращение) «гибридная сила», ускорение роста и увеличение размеров, повышение жизнестойкости и плодовитости гибридов первого поколения при различных скрещиваниях как животных, так и растений. Во втором и … Большая советская энциклопедия
Книги
- Социальная психология детства. Учебник , В. В. Абраменкова. Учебник посвящен анализу феномена детства в истории культуры и современности в контексте междисциплинарного направления социальной психологии детства. Авторомпрослежена эволюция важнейших…
Гетерозис (от греч. heteroiosis - изменение, превращение), «гибридная сила», понимают превосходство потомства I поколения над родительскими формами по жизнеспособности, выносливости, продуктивности,увеличение мощности возникающее при скрещивании разных растений. Во втором и последующих поколениях Гетерозис обычно затухает. Различают истинный Гетерозис - способность гибридов оставлять большое число плодовитых потомков, и гигантизм - увеличение всего гибридного организма или отдельных его частей.
Понятие о гетерозисе как проявлении «гибридной силы» было введено в науку американским генетиком В. Шеллом в 1914 г. Впервые явление гибридной силы наблюдал Ч. Дарвин у кукурузы. В его опытах у этой культуры снижалась продуктивность и уменьшалась высота растений в результате самоопыления, усиливались эти признаки при перекрестном опылении. Повышенную мощность растений, получаемых в результате скрещивания, Ч. Дарвин связывал с наследственными различиями родительских гамет.
Гетерозис в природе -- очень древнее явление. Он непосредственно связан с возникновением и совершенствованием в процессе эволюции способа перекрестного опыления. Естественный отбор на протяжении многих веков создавал многочисленные ограничения для гомозиготности и столь же многочисленные приспособления для осуществления гетерозиготности.
Гетерозис у гибридов проявляется в повышении роста, более интенсивном обмене веществ и большей урожайности. Повышенная урожайность гетерозисных гибридов -- главное их преимущество. Прибавка урожая у гибридов первого поколения всех сельскохозяйственных культур составляет в среднем 15--30 %, при этом нередко повышается их скороспелость. Например, у помидоров гетерознсные гибриды начинают плодоносить ка 10-- 12 дней раньше и превосходят по урожайности исходные родительские сорта на 45--50 %. В Болгарии все площади этой культуры заняты гетерозисными гибридами. Используя гетерозис, можно значительно увеличить производство сельскохозяйственной продукции.
При гетерозисе не обязательно происходит усиление всех свойств и признаков растений. По одним из них он может проявляться сильнее, чем по другим, а по некоторым отсутствовать.
Гетерозис наблюдается при скрещиваниях между сортами, а также между отдаленными в генетическом и экологическом отношении видами и формами. Наиболее же сильно он проявляется и поддается управлению при скрещивании самоопыленных линий.
Одна из характерных особенностей гетерозиса -- наибольшее проявление его у гибридов первого поколения, резкое снижение во втором поколении и дальнейшее затухание гибридной мощности растений в последующих поколениях. Это связано с уменьшением числа гетерозиготных особей. Например, если при скрещивании двух самоопыленных линий ААвв и ааВВ в первом поколении будет 100 % гетерозиготных растений, то во втором поколении их количество уменьшится в 2 раза, а в третьем -- в 4 раза и т. д.
И. В. Мичурин неоднократно указывал на преимущества сеянцев первой генерации и категорически возражал против использования в работе гибридов второй и третьей генерации, поскольку только у сеянцев первого гибридного поколения, обладавших вследствие гетерозиготности родительских сортов большим разнообразием признаков и свойств, гетерозис закрепляется при дальнейшем вегетативном размножении.
Важнейшее отличие гетерозисных гибридов от обычных гибридных сортов состоит в том, что их используют в производстве лишь в первом поколении и поэтому получают ежегодно.
Типы гетерозисных гибридов.
Различают следующие типы гибридов производственного использования: Простые(межсортовые, межлинейные, сортолинейные) и сложные(трехлинейные, четырехлинейные, синтетические). Межсортовые (редко).
При межсортовой гибридизации семена обоих компонентов высевают на участке гибридизации, где происходит их переопыление. Полученные семена передают в производство. У культур, у которых кастрация цветков не вызывает особых затруднений, все растения материнской формы кастрируют. У культур, у которых кастрация сопряжена с трудностями (свекла, подсолнечник, сорго и т. д.), происходит свободное переопыление родительских форм. В этом случае получают смесь, в которой будут и гибридные, и негибридные семена. Гетерозис здесь, естественно, ниже. Трудоемкость получения семян и их стоимость довольно высоки, а прибавки урожая небольшие -- до 10%. Поэтому межсортовые гибриды не получили широкого распространения. Только по сахарной свекле они занимают значительные площади.
Более эффективны сортолинейные гибриды. Самоопыленные линии (или инцухт-линии) получают у перекрестноопыляющихся культур путем принудительного самоопыления растений в течение 5--7 лет. С каждым годом самоопыления гомозиготность линий возрастает, а урожай в силу депрессии снижается, часто падая до 30--50% по сравнению с сходной формой. Увеличивается генетическая и морфологическая дифференциация линий. Отбирают линии с хорошо выраженными ценными хозяйственными и биологическими признаками: устойчивые к полеганию, осыпанию, с хорошей облиственностью, холодостойкие, засухоустойчивые, иммунные, с хорошими показателями качества и т. д.
Наиболее высокую прибавку урожая дают гибриды, полученные с участием самоопыленных линий. Поэтому в производстве распространены главным образом межлинейные и сортолинейные гибриды кукурузы, а межсортовые полностью вытеснены как менее урожайные..У кукурузы лучшие межлинейные гибриды по продуктивности превышают на 30--40% свободноопыляющиеся сорта. Таким образом, эти гибриды отличаются высокой гомогенностью (однообразностью) и высоким гетерозисом. Но они, как правило, очень дорогостоящие. Снизить высокую стоимость при производстве гибридных семян простых межлинейных гибридов можно: путем удачного выбора исходного материала возможно уменьшить инцухт-депрессию линий и тем самым повысить их урожайность или для этой цели используют так называемые сестринские скрещивания.При этом для каждой предназначенной для скрещивания линии А и В создают по одной сестринской линии А и В. Перед скрещиванием линий А и В проводят скрещивание соответствующих сестринских линий, чтобы значительно снизить инцухт-депрессию.
Как правило, двойные гибриды имеют почти такую же урожайность, как и простые, но уступают им по выравненности. Однако они часто характеризуются лучшей приспособляемостью к окружающей среде. В мировом производстве гибридной кукурузы двойные гибриды играли большую роль, чем простые межлинейные гибриды. Однако удельный вес простых гибридов кукурузы в производстве постепенно увеличивается.При использовании гибридов в производстве требуются постоянные закупки семян. Однако создание так называемых гибридных популяций позволяет повторно размножать гибридные семена.Гибридные популяции получают путем механического смешивания ряда самоопыленных линий и простых гибридов и свободного их переопыления. Cнижение уровня гетерозиса зависит от количества родительских форм (линий), участвующих в создании гибрида. В связи с этим гибридные популяции, представляющие смесь нескольких линий, имеют высокую урожайность не только в первом поколении после свободного переопыления смеси линий: в последующих поколениях гетерозис у них снижается в меньшей степени, чем у обычных гибридов. Таким образом, возможно повторное размножение гибридных семян. Недостаток гибридных популяций -- более низкая их урожайность по сравнению с двойными межлинейными или простыми гибридами.
Гетерозис , или гибридная мощность (сила) – это явление превосходства гибридов 1-го поколения по сравнению с исходными родительскими формами. Оно проявляется по многим признакам при скрещивании разных видов, рас, пород животных и сортов растений, а также инбредных линий.
Еще в середине XVIII в И. Кельрейтер , академик Российской академии, знаменитый ботаник, обратил внимание на тот факт, что в отдельных случаях при скрещивании растений гибриды первого поколения значительно мощнее своих родительских форм. Затем Ч. Дарвин сделал заключение, что гибридизация во многих случаях сопровождается более мощным развитием гибридных организмов.
Термин «гетерозис » ввел в 1914 г . американский генетик и кукурузовод Дж. Шелл .
Хотя эффект гетерозиса известен с древнейших времен, его природа до начала XX в. была неясна. Глубокий научный анализ явления гетерозиса стал возможен только после открытия основных генетических закономерностей.
Межлинейные гибриды кукурузы . В начале XX века Г. Шелл показал, что при скрещивании некоторых инбредных линий кукурузы получаются дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта.
Сейчас посев гибридными семенами стал основным приемом производства кукурузы . Для получения гибридных семян сначала создают большое количество инбредных линий из лучших сортов, отвечающих требованиям данного климатического района. Инбредная линия создается в течение 5 – 7 лет путем самоопыления . При отборе линий оцениваются качества, которые необходимо получить у будущего гибридного потомства. Значительная часть линий (около 99%) бракуется из-за тех или иных отрицательных свойств.
Создание большого количества инбредных линий – необходимый этап работы для получения гетерозисных форм. Особи в пределах линии имеют сходные генотипы и является практически гомозиготными. Поэтому при скрещивании таких линий образуются одинаковые по генотипу гетерозиготные гибриды .
Полученные таким образом межлинейные гибриды первого поколения оценивают по эффекту гетерозиса, отбирают линии, дающие лучшие комбинации, и затем размножают их в больших масштабах для производства гибридных семян . Чем больше создано инбредных линий, тем вернее и скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств. Чтобы найти пару линий , дающих при скрещивании высокий эффект гетерозиса , необходимо проверить несколько тысяч гибридных комбинаций .
При получении гибридных семян для производственных целей исходные линии , дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы . Для обеспечения опыления между ними разработана схема производства гибридных семян с использованием цитоплазматической мужской стерильности , что позволило значительно сократить затраты труда на удаление метелок с растений материнской линии . Так получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестно опыляющихся растений. Бывают также тройные гибриды (рис. 226).
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов . Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис (рис. 147).
Тройные идвойные гибриды получают в два этапа: 1) получение простого гибрида; 2) использование простого гибрида в качестве материнской формы для тройного или двойного, в качестве отцовских форм которых выступают линия и простой гибрид соответственно.
Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом селекции. Лучшие результаты дает скрещивание инбредных линий , происходящих из различных сортов . Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов – А × В , а другой – от скрещивания линий других сортов – C × D , то двойной гибрид (А × В ) × ( C × D ) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, каждый из которых происходит от линий одного сорта. Совершенно ясно, что в большинстве случаев инбредные линии всегда будут иметь более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только исходные линии , но и сорта или породы , от которых произошли эти линии.
Применение цитоплазматической мужской стерильности. Возникает вопрос, как получить гибридные семена, например, у кукурузы, сахарной свеклы, риса, томатов, если в пределах одного растения или даже одного цветка расположены женские и мужские элементы системы размножения и всегда присутствует возможность самоопыления. В этих случаях избежать процесса самоопыления можно только двумя путями: на материнских формах удалить вручную мужские элементы цветка, продуцирующие пыльцу; сделать мужские соцветия стерильными . Первый путь очень трудоемок, поэтому генетики начали поиск систем, определяющих мужскую стерильность растений.
В 1929 г. ученик Н. И. Вавилова отечественный селекционер и генетик Михаил Иванович Хаджинов нашел в посевах кукурузы растения с мужской стерильностью, которые ничем не отличались от нормальных, только были полностью стерильными, т. е. не продуцировали пыльцу. Это явление – цитоплазматическая мужская стерильность (ЦМС) – было изучено и широко использовано для получения гибридных семян у кукурузы, а затем и у многих других видов.
Схема использования ЦМС в селекции разработана в 30-х годах М. Родсом . Было установлено, что только взаимодействие особого типа цитоплазмы (S) и рецессивных генов ядра (rf) обусловливает мужскую стерильность (рис. 227, 228).
В практике используют лишь гибридные семена первого поколения от скрещивания двух линий, простого гибрида и линии или двух простых гибридов. Второе и последующие поколения в производственных посевах не используются, так как гибриды расщепляются на исходные формы и эффект гетерозиса исчезает. В связи с этим при использовании гетерозиса у растений организовано семеноводство в специальных хозяйствах, где получают только семена первого поколения и продают их хозяйствам, фермерам и т. д. Так как урожайность гетерозисных гибридов значительно (на 20-30%) выше исходных сортов, то затраты на семеноводство гибридных семян с лихвой окупаются. Внедрение гетерозисных гибридов кукурузы, по оценке американских специалистов, принесло чистый доход, в сотни миллиардов долларов. Гетерозис используется также при выращивании сахарной свеклы, риса, томатов и других видов.
Аналогичным образом получают гибриды и у животных. В настоящее время в птицеводстве и в свиноводстве широко используется скрещивание инбредных линий, происходящих из одной или разных пород .
Факторы , влияющие на гетерозис. Проявление гетерозиса у гибрида зависит:
1) от генотипа;
2) от свойств цитоплазмы , в результате чего реципрокные скрещивания дают разный эффект . Например, от скрещивания ♀ лошадь × ♂ осел получается высокогетерозисный гибрид мул – долговечный, выносливый и сильный. Реципрокная комбинация дает лошака , у которого гетерозис полностью отсутствует .
3) от стадии онтогенеза. Гетерозис в онтогенезе реализуется неравномерно . На одних стадиях онтогенеза проявляется гетерозис по одним признакам, на других – по другим. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть , например, в отношении устойчивости к неблагоприятной температуре . Гетерозис по этому свойству может проявиться позднее .
4) Сильное влияние на проявление гетерозиса оказывают также факторы среды .
Возможные механизмы гетерозиса. В настоящее время имеются 4 гипотезы , объясняющие возникновение гетерозиса:
1. Гипотеза гетерозиготности гибридов Дж. Шелла и Е. Иста (1908 г.). Как уже говорилось, при скрещивании гомозиготных инбредных линий гибриды первого поколения гетерозиготны по многим генам . При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей.
Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена (ааВВ) , а вторая – другого гена (ААbb) . Каждая из этих рецессивных аллелей в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии.
При скрещивании линий aaBB × AAbb у гибрида объединяются доминантные аллели обоих генов (AaBb ) . Гибриды F 1 проявят при этом по указанным генам не только гетерозис , но и единообразие .
ВF 2 число особей с двумя доминантными генами в гетерозиготном состоянии (AaBb) будет лишь 4 / 16 , поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается . В силу этих причин гетерозис в последующих поколениях затухает .
Серьезным препятствием для этой гипотезы является тот факт, что далеко не все межлинейные гетерозиготные гибриды проявляют гетерозис , т. е. гетерозиготность не всегда связана с гетерозисом.
2. Гипотеза доминирования Д. Джонса (1918 г.) исходит из того факта, что доминантные аллели дикого типа чаще, чем рецессивные, оказывают благоприятное действие независимо от того, в каком состоянии - гомозиготном или гетерозиготном – они находятся. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Иначе говоря, эта гипотеза исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием .
Проиллюстрируем это следующим примером :
P: AAbbCCdd × aaBBccDD
Если у скрещиваемых форм имеется всего по два доминантных благоприятно действующих гена, то у гибрида их четыре, независимо от того, в гомозиготном или гетерозиготном состоянии они находятся. Это, по мнению сторонников этой гипотезы, и определяет гетерозис гибрида, т. е. его преимущества перед исходными формами.
И эта гипотеза вступает в противоречие с рядом фактов. Так, при инбридинге идет гомозиготизация . Следовательно, согласно этой гипотезе, следовало ожидать появления в результате инбридинга гетерозисных форм , имеющих набор доминантных генов в гомозиготном состоянии, однако это не имеет места.
3. Гипотеза сверхдоминирования Шелла и Иста исходит из того, что гетерозиготное состояние имеет превосходство над гомозиготным (АА < Аа > аа). Предполагается, что сочетание в гетерозиготе аллелейдикого типа и мутантного каким-то образом усиливает действие доминантного гена и в связи с этим вызывает максимальное накопление специфических веществ , синтез которых контролируется этим геном.
4. Гипотеза компенсационного комплекса генов В. А. Струнникова . Ее суть сводится к тому, что при возникновении мутаций сильно понижающих жизнеспособность и продуктивность организма, так называемых полулеталей , при отборе у гомозигот формируется компенсационный комплекс генов , в значительной степени нейтрализующий вредное действие мутаций. Если затем такую мутантную форму скрестить с нормальной без мутации и тем самым перевести мутацию в гетерозиготное состояние , т. е. нейтрализовать ее действие нормальным аллелем, то сложившийся по отношению к мутации компенсационный комплекс в гибридном организме будет «работать» на гетерозис .
Пути закрепления гетерозиса. Основной задачей использования гетерозиса в селекции является закрепление его , т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида . Решение этой задачи возможно следующими путями.
1. Закрепить гетерозис удается там, где возможно вегетативное размножение гибридов – клубнями, луковицами, черенками. Так сохраняется положительный эффект гетерозиса у картофеля.
2. Важным подходом к закреплению гетерозиса является кратное увеличение наборов хромосом – полиплоидия .
3. В естественных условиях гетерозис закрепляется при возникновении полиморфизма по инверсиям .
4. У животных, для которых все эти пути закрепления гетерозиса исключены, используют переменное скрещивание , т. е. регулярное скрещивание гибридов попеременно с одной и с другой исходными формами.
5. Аналогично опытам Гердона возможна пересадка ядер гетерозисных животных в энуклеированные яйцеклетки, т. е. клонирование .
В селекции животных и растений особое место занимает явление гибридной мощности, или гетерозиса , которое заключается в следующем.
При скрещивании разных рас, пород животных и сортов растений, а также инбредных линий гибриды F 1 по ряду признаков и свойств часто превосходят исходные родительские организмы. Скрещивание гибридов между собой ведет к затуханию этого эффекта в следующих поколениях. В настоящее время гетерозис установлен для всех изученных видов.
Хотя эффект гетерозиса известен с древнейших времен, его природа до сих пор остается мало изученной. Первое толкование биологического значения гетерозиса в эволюции животных и растений и попытка объяснить механизм этого явления принадлежали Ч. Дарвину. Он систематизировал большое количество фактов и сам ставил опыты по изучению гетерозиса. По мнению Ч. Дарвина, гетерозис служит одной из причин биологической полезности скрещивания в эволюции видов. Перекрестное оплодотворение поддерживается естественным отбором именно потому, что оно служит механизмом сохранения наибольшей гетерозисности,
Глубокий научный анализ явления гетерозиса стал возможен только с начала XX в. после открытия основных генетических закономерностей. При рассмотрении гетерозиса мы будем касаться возникновения его при скрещивании преимущественно инбредированных линий одного вида, поскольку в этом случае легче выяснить его генетический механизм.
С начала нашего столетия на кукурузе стали проводить систематическое исследование скрещиваний между инбредными линиями. При этом Г. Шеллом было показано, что скрещивание некоторых линий дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта. Приведены опытные данные, показывающие низкую урожайность инбредных линий, значительное повышение урожайности в F 1 и снижение в F 2 при самоопылении растений F 1 .
За последние 30 лет в ряде стран, в том числе и в нашей, посев гибридными семенами стал основным приемом производства кукурузы как на зерно, так и для силоса. Для получения гибридных семян сначала создают инбредные линии из лучших сортов, отвечающих требованиям данного климатического района (инбредная линия создается в течение 5-6 лет путем самоопыления). При отборе линий оценивают их качества и свойства, которые необходимо получить у будущего гибридного организма. Инбридинг в линиях не может быть эффективным, если он не сопровождается отбором.
Создав большое число инбредных линий, приступают к скрещиванию между ними. Межлинейные гибриды первого поколения оценивают по эффекту гетерозиса ; исходя из этого показателя отбирают линии с лучшей комбинационной ценностью и затем размножают их в больших масштабах для производства гибридных семян. Работа по созданию инбредных линий и оценке их комбинационной ценности постоянно ведется в селекционных учреждениях. Чем больше создано ценных линий, тем вернее й скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств.
При получении гибридных семян для производственных целей исходные линии, дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы. Чтобы обеспечить опыление между ними, с материнских растений удаляют мужские соцветия (метелки). Теперь разработана новая схема производства гибридных семян с использованием цитоплазматической мужской стерильности, что позволило значительно сократить труд на удаление метелок у растений материнской линии. Таким путем получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестноопыляющихся растений.
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов. Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис.
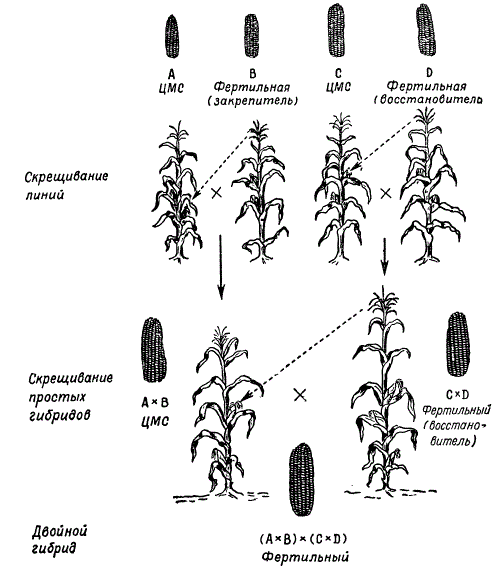
По данным М. И. Хаджинова и Г. С. Галеева, результаты оценки продуктивности разных гибридов - межсортовых, сортолинейных и двойных межлинейных - показывают, что наиболее продуктивны двойные межлинейные гибриды.

Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом их селекции. Лучшие результаты дает скрещивание линий, происходящих из различных сортов. Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов А X В, а другой - от скрещивания линий других сортов С х D, то двойной гибрид (А X В) х (С X D) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, происходящих от линий одного сорта: (А х А 1 х (А 2 х А 3) или (В х B 1) х (В 2 х В 3).
Для успешного развития гибридного семеноводства необходимо исследовать, во-первых, как долго следует вести инбридинг, чтобы получить гомозиготные линии, и, во-вторых, разработать методы более быстрой оценки их комбинационной ценности.
Все, что было сказано в отношении кукурузы, по-видимому, является общим для получения гибридов у других высших перекрестноопыляющихся растений, а также у животных. В настоящее время в птицеводстве и в свиноводстве ряда стран широко используется скрещивание инбредных линий, происходящих из одной или разных пород. Следует особо подчеркнуть, что широкое использование гибридов в животноводстве возможно только при высоком уровне племенной работы и наличии ценных пород, то совершенно ясно, что в большинстве случаев инбредные линии в среднем будут иметь всегда более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только родителей (линии), но и сорта, или породы, от которых произошли эти линии.
Как мы знаем, гены детерминируют свойства организма на всех стадиях онтогенеза начиная с момента оплодотворения. Гены ядра ооцита способны детерминировать свойства цитоплазмы яйцеклетки еще до оплодотворения. Характер реализации генотипа изменяется в зависимости от свойств цитоплазмы зиготы. Таким образом, проявление гетерозиса у гибрида зависит также и от свойств цитоплазмы. Роль цитоплазмы в определении гетерозиса доказывается следующим образом. При реципрокном скрещивании двух линий А X В и В х А гетерозис по одним и тем же свойствам нередко проявляется у гибридов только одного из скрещиваний и не проявляется у гибридов другого.
Проявление гетерозиса обусловлено индивидуальным развитием гибрида. В онтогенезе он реализуется неравномерно. На одних стадиях онтогенеза проявляется гетерозис по одним признакам, а на других - по другим или только по некоторым. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть, например, в отношении устойчивости к неблагоприятной температуре. Гетерозис по этому свойству может проявляться позднее.
Сильное влияние на проявление гетерозиса оказывают также факторы среды, в которых развивается гибридный организм. Есть основание считать, что гетерозис по жизнеспособности и устойчивости к заболеваниям лучше выявляется при неблагоприятных для развития условиях среды. Еще Ч. Дарвином было высказано предположение, что гетерозис у гибридов обусловлен более широкой нормой приспособительных реакций. Исследования последних лет показали, что гибриды F 1 в сравнении с инбредными линиями характеризуются большим разнообразием процессов обмена, участием большего числа различных метаболитов, ростовых веществ и энзимов. Гетерозис проявляется не только в целом организме, но и на клеточном уровне.
Выяснение генетических механизмов гетерозиса остается нерешенной проблемой. В настоящее время имеются три гипотезы, пытающиеся объяснить возникновение гетерозиса:
- гетерозиготное состояние по многим генам,
- взаимодействие доминантных благоприятных генов,
- сверхдоминирование - гетерозигота превосходит гомозиготы.
Как мы уже говорили, при скрещивании гомозиготных инбридированных линий гибриды первого поколения по многим генам приобретают гетерозиготное состояние. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей. Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена ааВВ, а вторая - другого гена ААbb. Каждая из этих рецессивных аллелей генов в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии. При скрещивании линий ааВВ X ААbb у гибрида объединяются доминантные аллели обоих генов (АаВb). Гибриды F 1 проявят при этом по указанным генам не только гетерозис, но и единообразие. В F 2 число особей с двумя доминантными генами в гетерозиготном состоянии будет лишь 4/16, поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается. В силу этих причин гетерозис в последующих поколениях затухает. Такова схема одной из генетических гипотез гетерозиса.
Мы привели пример с двумя генами, но физиологические свойства определяются огромным числом генов. Кроме того, неверно считать, что доминантные аллели всегда положительно, а рецессивные отрицательно влияют на жизнеспособность организма и его продуктивность. Доминантные аллели дикого типа чаще оказывают благоприятное действие, чем рецессивные. Это можно объяснить тем, что неблагоприятные доминантные мутации подвергаются более жесткому отбору - они уже в зиготе и на эмбриональных стадиях подвергаются элиминации, и отбором сохраняются лишь благоприятные. И так как само свойство доминирования гена эволюционирует под контролем отбора, то аллели дикого типа и оказываются более благоприятными для организма. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Если бы гетерозис обусловливался простым набором доминантных аллелей, имеющихся в популяции, то этот набор было бы легко составить путем ряда скрещиваний и получить гетерозисные сочетания. Весьма возможно, что часть пород и сортов выводилась именно путем скрещивания и комбинаций линий, имеющих набор благоприятных доминантных аллелей. Но до сих пор не удавалось закрепить гетерозис в гибридной комбинации F 1 , т. е. получить не расщепляющиеся в F 2 формы.
Д. Джонсом еще в 1917 г. было предложено дополнение к изложенной гипотезе. Согласно Д. Джонсу, разные гены, дающие в сочетании гетерозис, находятся в пределах одной группы сцеплений: например, AbcdE - в одной, а соответственно aBCDe - другой гомологичной хромосоме. Именно это обстоятельство, с точки зрения Д. Джонса, затрудняет отбор в F 2 полностью гомозиготных форм по благоприятным доминантным генам, дающим сочетании гетерозис. Для осуществления такого сочетания доминантных аллелей необходимо, чтобы в этой паре хромосом AbcdE//aBCDe произошел минимум двойной кроссинговер, который привел бы к возникновению хромосомы с одними доминантными аллелями ABCDE. Малая вероятность одновременного осуществления такого события в нескольких хромосомах и является, по мнению Д. Джонса, причиной, затрудняющей закрепление гетерозиса. Гибрид же F 1 содержит все эти доминантные аллели, и поэтому у него наблюдается гетерозис.
Такова схема объяснения гетерозиса согласно второй гипотезе - гипотезе взаимодействия доминантных благоприятных генов, которую называют иногда также гипотезой набора кумулятивных доминантных аллелей, а сам гетерозис при этом - мутационным. Эта гипотеза в целом исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием.
Третья гипотеза исходит из того, что гетерозиготное состояние аллелей имеет превосходство над гомозиготным (АА < Aa > аа). Здесь можно допустить благоприятное трансположение аллелей дикого типа и мутантных аллелей, которое каким-то образом усиливает эффект действия генов. Указанное объяснение гетерозиса называют гипотезой сверхдоминирования.
Ни одна из трех гипотез не может считаться единственно правильной. Сейчас преждевременно отдавать предпочтение той или другой. Вероятно, они все окажутся правильными, но для разных случаев. Наверное, каждый из механизмов, предусматриваемых этими гипотезами, играет роль в определении гибридной мощности. Гетерозис является сложным явлением как по механизму возникновения, так и по проявлению его в онтогенезе.
Очевидно, окончательный вывод о генетических механизмах гетерозиса можно будет сделать лишь после того, как раскроется картина взаимодействия генов в генетической системе на биохимическом и молекулярном уровнях. Как мы уже знаем, подавление проявления мутаций с помощью супрессоров позволяет исправлять нарушения в действии гена и его аллелей. Возможно, что явление гетерозиса есть суммарный результат работы не основных генов, определяющих развитие признаков организма, а набора супрессирующих генов в генотипе. Важным моментом изучения гетерозиса становится исследование плазменных отношений.
Основной задачей использования гетерозиса в селекции является закрепление его, т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида. Решение этой задачи мыслится в нескольких аспектах: во-первых, закрепление гетерозиса путем перевода гибридного организма с полового размножения на апомиктическое, что, по-видимому, возможно для некоторых растений; во-вторых, закрепление гетерозиса путем перевода диплоидного гибрида, проявляющего гетерозис, в полиплоидное состояние. В этом случае гетерозиготная комбинация генов будет сохраняться дольше.
У вегетативно размножающихся растений поддержание ценных гибридных комбинаций, полученных половым путем, осуществляется вегетативным размножением (черенками, прививками, клубнями и т. д.).
Существует ряд других путей поддержания гетерозиса в ряду поколений, однако все они еще недостаточно разработаны и проверена.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Гетерозис - это свойство помесей и гибридов первого поколения (Fi) превосходить по биологическим и хозяйственно-полезным признакам исходные родительские формы.
Впервые явление гетерозиса было описано И. Кёльрейтером , работавшим в Петербургской академии наук, на примере межвидового гибрида, который он получил в 1760 г., скрестив два разных вида махорки . Этот растительный гибрид оказался стерильным наподобие мула и автор назвал его «первым растительным мулом».
Научный термин «гетерозис» появился гораздо позже. Его предложил американский исследователь Дж. Шелл в 1914 г. для обозначения мощности гибридов (эффекта скрещивания), и с тех пор он прочно вошел в научную литературу как синоним старого названия «гибридная сила».
В настоящее время твердо установлено, что гетерозис проявляется не только при разведении животных и птиц, но и при селекции растений, а также микроорганизмов. Следовательно, гетерозис - явление общебиологическое.
Каковы причины возникновения гетерозиса? На этот счет существует несколько точек зрения в виде отдельных самостоятельных гипотез.
Одна из первых изложена в фундаментальном труде Чарльза Дарвина «Действие перекрестного опыления и самоопыления в растительном мире», в котором он изложил вопросы гетерозиса («гибридной силы»).
Изучив опыт английских заводчиков по созданию новых пород сельскохозяйственных животных, Ч. Дарвин отметил, что родственное спаривание (инбридинг), которое применялось им для закрепления в потомстве желательных признаков выдающихся производителей, приводит, как и самоопыление у растений, к отрицательным последствиям - к депрессии, в то время как скрещивание повышает, как правило, жизнеспособность потомства за счет проявления эффекта скрещивания (гетерозиса).
Дарвин предположил, что в основе этих двух явлений - инбредной депрессии и гетерозиса - лежит одна и та же причина - степень различия половых элементов, объединяющихся в процессе оплодотворения.
Чем больше родительские формы, а следовательно, и их половые клетки различаются по своим биологическим особенностям, тем сильнее в потомстве проявляется гетерозис, и наоборот, отсутствие таких различий при близкородственном длительном спаривании приводит к нибредной депресии.
Исходя из этих соображений, Ч. Дарвин провозгласил «великий закон природы», по которому, сточки зрения эволюции вида, скрещивание всегда полезно, а родственное спаривание (инбридинг - у животных, самоопыление - у растений) - вредно.
Гипотеза доминирования (Джонс, 1972). В основе этой гипотезы лежит идея благоприятного действия доминантных генов - гетерозис проявляется в результате взаимодействия при скрещивании благоприятных доминантных факторов, имеющихся у исходных родительских форм.
Предполагается, что при скрещивании происходит сочетание благоприятно действующих неаллельных доминантных генов и одновременное подавление ими действия различных вредных рецессивных аллелей, которые у разных линий, а тем более пород, находятся в разных локусах. При скрещивани доминантные аллели, внесенные одним родителем (линией), могут перекрывать рецессивные аллели, полученные гибридом от другой родительской формы (линии).
Проявление гетерозиса возможно также и за счет явления эпистаза, когда отдельные неаллельные гены (эпистатический ген) подавляют не только «свои» рецессивные, но и «чужие» доминантные гены (гипостатический ген).
Гипотеза доминирования общепризнана, однако, она не объясняет полностью все вопросы, возникающие в связи с проявлением гетерозиса. Так, если исходить из названной гипотезы, то теоретически следует ожидать, что при полигибридном скрещивании гетерозигота Аа будет в той или иной степени приближаться по продуктивности к гомозиготе АА - приближаться, но не превосходить ее.
Однако в практике давно установлено, что гетерозигота может превосходить по мощности не только рецессивную родительскую форму, но и доминантную, то есть обоих своих родителей. Это явление получило в генетике даже специальное название - сверхдоминирование, или моногибридный гетерозис.
Гипотеза гетерозиготности (сверхдоминирование). С точки зрения гипотезы гетерозиготности проявление гетерозиса объясняется, говоря современным языком, разнокачественностью членов одной и той же пары аллелей у гибридных организмов как результат скрещивания различающихся исходных родительских форм. Соединение при гибридизации разнокачественных гамет родителей уже само по себе стимулирует более быстрый рост гетерозиготных гибридов, их лучшее развитие и т. д. В результате гибрид по мощности превосходит исходные гомозиготные родительские формы как рецессивную, так и доминантную, что и обусловливает эффект сверхдоминирования. В то же время гомозиготность родителей оказывает угнетающее действие на жизнеспособность потомства, что выражается формулой Аа>АА>аа.
Предполагается, что у гетерозиготы оба аллеля одного локуса выполняют различные функции, взаимно дополняя друг друга в биохимическом процессе. При этом эффект гетерозиса будет тем выше, чем в большей степени аллели каждого локуса различаются функционально между собой, чем больше они дополняют друг друга.
Причины гетерозиса, указанные в этих гипотезах, возможно, действуют одновременно, однако их, по-видимому, не достаточно для всестороннего объяснения механизма возникновения гетерозиса как общебиологического явления. По этому поводу проф. М. В. Лобашов (1969) писал: «Трудно предположить, чтобы в основе столь сложного явления, как гетерозис, лежал единственно генетический механизм». Что же касается понимания самого механизма генного взаимодействия при гетерозисе, то по современным воззрениям различие между обеими гипотезами незначительно или его совсем нет.
Гипотеза генетического баланса (академик ВАСХНИЛ Н. В. Турбин, 1961. С точки зрения этой гипотезы явление гетерозиса нельзя объяснить действием одной какой-либо генетической причины - это суммарный эффект.
Гипотеза генетического баланса , принимая в обеих чертах отдельные положения ранее высказанных гипотез, больше внимания, однако, уделяет взаимовлиянию неаллельных генов, физическим и биохимическим факторам, а также внешней среде вообще, условиям выращивания гибридов в частности . Особое внимание придается цитоплазматическому влиянию. Предполагается, что плазматические различия между гаметами должны стимулировать жизненные процессы у гибридного организма. Сбалансированность систем генов делает популяции наиболее приспособительными и продуктивными в конкретных условиях среды.
Следует отметить, что в последние годы все большее значение приобретает также биохимическая теория гетерозиса , согласно которой скрещивание приводит к увеличению гетерозиготности по мутациям, регулирующим синтез белка - отсюда проявление гетерозиса происходит за счет обогащения биохимических процессов в клетках и тканях гибридного организма.
Значение изложенных выше гипотез неоспоримо, однако ни одна из них не может быть пока признанной в качестве общепринятой теории гетерозиса. Возможно, прав известный генетик Ф. Хатт, в своем высказывании: «Гетерозис все еще представляет одну из самых больших загадок генетики».
Что же представляет собой гетерозис как генетическое явление? Каковы формы проявления гетерозиса?
Гетерозис - это совокупность явлений, связанных с повышенной жизнеспособностью помесей (гибридов), которая, как установлено, проявляется уже на ранних стадиях развития (онтогенеза) . У эмбрионов гибридных кур, например, еще в эмбриональном развитии усиливаются обменные процессы, ускоряется их развитие, в результате чего вывод и качество суточного молодняка бывают выше по сравнению с теми же показателями линейных кур (Злочевская, 1968).
Гетерозис - явление неустойчивое (кратковременное), оно наиболее ярко (четко) проявляется лишь в первом поколении (F 1) скрещивания. Помесные (гибридные) животные при их дальнейшем разведении не дают подобных себе гетерозисных потомков, онине остаются «константными» по гетерозису. Поэтому их не оставляют на племя, а реализуют на мясо. Следовательно, гетерозис нельзя закрепить наследственно, его нужно всякий раз получать заново.
В отдельных случаях гетерозис можно поддерживать на относительно высоком уровне и в последующих поколениях, но в таких случаях используются специальные методы - переменное скрещивание и др. Считается, что угасание гетерозиса в последующих поколениях гибридов - результат рекомбинационных утерь .
Формы проявления гетерозиса бывают различными. Обычно при скрещивании двух пород (А и В) уровень продуктивности помесного (АВ) потомства равен средней показателей продуктивности исходных пород В таких случаях говорят о гипотетическом (вероятном) гетерозисе.
![]()
Нередко продуктивность помесных (F 1) животных оказывается значительно выше средней продуктивности родителей, а иногда она превышает показатели лучшей из родительских форм - абсолютный (истинный) гетерозис.
![]()
В других случаях, однако, продуктивность помесей превышает показатели лишь одного из родителей, худшего - относительный гетерозис:
![]()
где: Пг - признак гибрида; Пл - признак лучшей породы; Пм - признак материнской породы; По - признак отцовской породы.
Конечно, из чисто практических соображений эффект гетерозиса представляет наибольший интерес лишь в том случае, когда гибридное потомство превышает по своей общей хозяйственно полезной ценности лучшего из родителей. Только в таких случаях скрещивание имеет экономический смысл. Поэтому селекционеры-практики под гетерозисом понимают свойство гибридов (F 1) превосходить по определенным признакам лучшую из родительских форм.
Некоторые ученые, учитывая специфику форм проявления гетерозиса, выделяют самостоятельные его типы:
репродуктивный гетерозис - более высокая общая продуктивность животных, связанная с повышением плодовитости (фертильности) и более мощным развитием их репродуктивных органов;
соматический гетерозис - более сильное развитие вегетативных частей (у растений), органов и частей тела (у животных);
адаптивный гетерозис - повышенная жизнеспособность животных, их лучшая приспособляемость.
У мулов, например, сильно выражен соматический гетерозис, то есть большая живая масса; выше тяговое усилие; повышенное долголетие; особая выносливость; но в то же время репродуктивная система недоразвита. Как правило, они бесплодны. Сказанное выше - это пример частного гетерозиса (мощное развитие касается не всего организма животного, а лишь его отдельных признаков) в отличие от общего гетерозиса, когда имеет место развитие общей массы тела животного, повышение метаболических процессов в организме в целом, что обеспечивает повышение его продуктивности (Красота, 1979).
Следует отметить, что гетерозис проявляется у помесей и гибридов - межвидовых, межпородных, межлинейных - по ограниченному числу признаков. Он никогда не проявляется по сумме всех родительских признаков. Помеси (гибриды) превосходят своих родителей не по всем показателям продуктивности, не по всем признакам, не по их сумме, а лишь частично, по отдельным признакам (или группе признаков) или даже по отдельно взятому признаку.
Метисные куры первого поколения, полученные от скрещивания петухов мясных пород с курами яйценоских (легких) пород, могут превосходить исходные родительские формы по яйценоскости, но по живой массе они занимают промежуточное положение.
Отсюда под гетерозисом следует понимать превосходство потомства - помесей или гибридов - над родительскими формами не по всем, а лишь по определенным, конкретным признакам.
Литература
1. Вавилов Н. И. Центры происхождения культурных растений.- Тр. по прикладной ботанике и селекции, 1926, т. XVI, с. 5-138.
2. Гершензон С. М. Основы современной генетики.- Киев, 1979.- 506 с.
3. Лобашев М. Е., Ватти К. В., Тихомирова М. М. Генетика с основами селекции.- М., 1979.- 304 с.
4. Рокицкий П. Ф. Некоторые этапы развития генетики животных в СССР и ее связи с селекцией.- В сб.: Генетические основы селекции животных. М., 1969, с. 9-25.
Рекомендуем также
 Всё о гарантийном ремонте товара
Всё о гарантийном ремонте товара
 Относительная величина выполнения плана
Относительная величина выполнения плана
 Защита прав работников в России: всё, что нужно знать руководителю и подчиненному
Защита прав работников в России: всё, что нужно знать руководителю и подчиненному
 “Ошибочно” присланное письмо о переводе средств На майл пришло письмо о переводе денег
“Ошибочно” присланное письмо о переводе средств На майл пришло письмо о переводе денег
 Увольнение сотрудника по болезни или состоянию здоровья – правила оформления и выплаты
Увольнение сотрудника по болезни или состоянию здоровья – правила оформления и выплаты
 Стив Шапиро. Проживая Америку. Стив Шапиро: Проживая Америку Проживая америку выставка фотографий стива шапиро
Стив Шапиро. Проживая Америку. Стив Шапиро: Проживая Америку Проживая америку выставка фотографий стива шапиро